



���Ӷ�ũ�˾��鵼��ֲ���Ŵ�ת����ϵ��Ӱ��

����ֲ��ݣ����ޣ��������������壬������*
���Ӷ�ũ�˾��鵼��ֲ���Ŵ�ת����ϵ��Ӱ��
�1,3,4���ֲ���1,3,4������2,3,4��������1,3,4��������1,3,4��������1,3,4*
1. ����ũ�ִ�ѧ��ѧԺ������ ���� 350002��
2. ����ũ�ִ�ѧ��Ϫ��ѧԺ������ Ȫ�� 362400��
3. ����ʡ���ҵ���̼����о����ģ����� ���� 350002��
4. ��Ͽ������ɫ���ﰲȫ����ʡ������Эͬ�������ģ����� ���� 350002
Ϊ�о���ũ�˾��鵼�IJ����Ŵ���ϵ�У����Ӷ�ũ�˾���ȾЧ�ʵ�Ӱ�죬��LBA4404��EHA105��ATCC15834��K599 4��ũ�˾�����Ϊ�о����������ڲ�ͬŨ�Ȳ��Ӵ����µ��ͷ���������Ĥ�����ʡ������ԡ��ͻ������Լ��Ŵ�ת�����졣�����ʾ��4��������ͷ���������ΪLBA4404��K599��EHA105��ATCC15834������EHA105��ATCC15834����Բ��ӽ�Ϊ���У�
ATCC15834���걻Ĥ����������ӵ�Ũ�ȡ�����ʱ��ʸ���أ�EHA105��600?mg��L-1���Ӵ���48?h���䱻Ĥ��������ͣ�
ATCC15834��EHA105���̲�Ҷ��ϸ���������������Ų���Ũ�ȵ����߶����ͣ��ھ����Ӵ���24?h��ATCC15834�к�EHA105�еı����ܵ��������ƣ����������Ũ�ȷ���������Գ�������ԣ�
�̲ݾ�ũ�˾���Һ�������ӣ���Ⱦ��˲ʱת��Ч�ʺ��ȶ�ת��Ч�ʾ��ܵ���ͬ�̶ȵ����ƣ���״���յ��ʴ�͡�1?000?mg��L-1���Ӵ�����ATCC15834�ķ����յ�����ͣ���Ϊ13.85%�������ʸߴ�33.85%���������������ӻή��ũ�˾������ͱ�Ĥ�����ʣ�����ͨ�����ͱ�����Ӱ���������ԣ����յ����̲�ת����ϵ��˲ʱת��Ч�ʺ��ȶ�ת��Ч�ʾ��ܵ���ͬ�̶ȵ����ơ�
���ӣ�
ũ�˾��鵼��
�����ԣ�
�ͻ���
�Ŵ�ת��
�����Dz�Ҷ�ж�������ʵ��ܳƣ�����Ҫ�ɷְ������鴼�ࣨ�Զ����������Ϊ��������ɫ���ࡢ��ͪ�ࡢ��ͪ����ͷ������[1-2]��������Ϊһ�ֹ��ס�ǿЧ���Ͷ��Ŀ���ҩ��Ը��������Ծ����������Ծ�����������������[3-4]�����о����������Ӷ�12��ϸ����Ⱥ������ϸ�������������ã����־�������Ũ�ȳ������[5-9]��
ũ�˾�����Ϊ��������ũ�˾�������������ϸ������ũ�˾��鵼���Ŵ�ת����ϵ�У������������ֲ���Ŵ�ת���о�����Ҫ�����á���Ũ�ȶ�������ʿ���Ϊ��ѧ������������ũ�˾���ֲ��ϸ���ƶ���������ũ�˾��������ϵͳ[10-12]�������С��[13]��ˮ��[14]�������Ŵ�ת���ʡ����ض�Ũ�ȷ�Χ�ڣ�����������ڲ����Ŵ�ת����ϵ�а����־��������ã���Ҫ��3��ԭ��1����ӻ���һ����Χ������ũ�˾����������ã���ֹũ�˾�����ֲ���˿ڣ�Ӱ��ũ�˾���ֲ��ϸ��֮���������[15-22]��
��2����ӻ�����ũ�˾���������������ʹ��������ܵ����ơ�vir���ױ������ȣ�
��3����������ʵ��������û����һ�����ɫ�ľۺ��������֯��������ֲ��ֻ�������[23]�����Ͽ����Dz���Ӱ�����ת��Ч�ʵ͵���Ҫԭ��
�ڲ������Ŵ�ת����ϵ�У�ǰ�˶�ũ�˾�����չ��������о����δ���[24]�о����֣�ũ�˾���������֯��ȡҺ��������������
�ڲ���Ҷ��ȡҺ�У�����ũ�˾�GV3101�ͷ���ũ�˾�ATCC15834�����ܵ�Ӱ�죬ʹ�á��ͻ�������ܵ��������ƣ���������������������Ҷ�еĶ����������ʡ�ë�����[25]�о����֣������ضԷ���ũ�˾�R1000���������á���÷�۵�[26]ͨ���о�A4��R12556��4��ũ�˾�����IJ��������ԣ�����������δ���Ӳ��ӵ��������ϣ�����36?h�ھ����ֶ�������������ͬŨ�Ȳ��Ӷ�ũ�˾�����������Ч�����������Բ��졣
���о�ѡȡ����ũ�˾���EHA105��LBA4404���ͷ���ũ�˾���K599��ATCC15834���������Ӷ�ũ�˾�����������Ӱ�죬������ѡȡ���жȸߵľ����һ���о����Ӷ��䱻Ĥ�����ԡ��������Լ���ػ����������Ӱ�죬̽�ֲ��Ӷ�ũ�˾��鵼���Ŵ�ת����ϵ�����û��ơ���ũ�˾��鵼�IJ����Ŵ�ת����ϵ�У����о�ּ��Ϊ����Ʒ�֣����˵IJ��Ӻ������������ѡ���ṩ�������ݣ�Ϊ������������ѧ�����ṩ���ۻ�����
1.1 �������
���ӣ����ȡ�99%���������Ϻ�����ѧ��������˾���̲�������Դ�ڸ���ũ�ִ�ѧ��Ͽ�����о�Ժ���������ģ�����������������������1/2 MS��������25�棬ÿ�����12?h������������6��8Ƭ��Ҷ��
ũ�˾�������ø���ũ�˾�EHA105��LBA4404������ũ�˾�K599��ATCC15834��
���о���EHA105��LBA4404��K599���˾�DH5Ϊ��ʵ���ұ��棬����ATCC15834�ɰ���ũҵ��ѧ�Խ������ṩ��˫Ԫ��������pBI121��Я��GUS����Ϊ��ʵ���ұ��棻
ԭ�˹�������������ΪpBBR1MCS2-Tac-EGFP��Я��eGFP��ǻ����������;�Ϊ����ù�أ������ں��Ϸ�������Ƽ�����˾��
YP��������������ĸ��ȡ��5?mg��L-1��������10?mg��L-1��NaCl 10?mg��L-1����֬12?mg��L-1��pH 7.0��121�����20?min��
YMB����������ĸ��ȡ��1?mg��L-1����¶��10?mg��L-1��K2HPO40.5?mg��L-1��MgSO4��7H2O 0.2?mg��L-1��NaCl 0.1?mg��L-1��pH 7.0��121�����20?min��
1.2 ���鷽��
1.2.1 ũ�˾��Ļ���Һ�Ʊ�
ũ�˾��������Ӧũ�˾������ں������ص�YP�����������У�����ƽ��Rifampicin��Rif������Ũ��Ϊ25?mg��L-1�������Ӧ�Ŀ���������Ũ�Ⱦ�Ϊ50?mg��L-1��28��������а�����2?d���������ֻ���ȡ����¡��YPҺ���������У�28�����ҡ����200?r��min-1�������ٴλ����Һ���ǣ�����Һǹȡ��1?000?��L��Һ������100?mL YPҺ��������������������OD600Ϊ0.6��0.8��
��Һ���Ʊ�������õ�ũ�˾���Һ����10?000����15?min��ȥ������Һ������ȾҺ��1/2 MS+0.20?g��L-1��ˮ���ҵ���+10?g��L-1����+0.5?g��L-1MES���������ֱ��������Ũ��Ϊ200��400��600��800?mg��L-1��1?000?mg��L-1�IJ��ӣ�����1����������ˮ������2�ú���Ӧ���ӵ���ȾҺ���棨����ũ�˾����������Ϊ50?mL�����¾���1?h��������Ⱦ�̲�Ҷ�̡�
1.2.2 ũ�˾������ʵIJⶨ
�ں��в�ͬŨ�Ȳ��ӵ�YMB�������м����õ�ũ�˾���Һ����ʼŨ�ȵ�OD600=0.05���ֱ���3��6��9��12��18��24��30��36��48��60?hȡ1?mL��Һ���Ժ���ӦŨ�Ȳ��ӵ�YMB������Ϊ�αȣ��ⶨ600?nm������ֵ�����Ϊ1?cm��������3���ظ�����¼���������ߣ�������������������Ӷ�ũ�˾����־��ʣ���ʽ���£�
���Ӷ�ũ�˾���������=��1��������OD600/������OD600����100%
1.2.3 pBBR1MCS2-Tac-mCherryԭ�˱������幹��
��pBBR1MCS2-Tac-EGFP������EcoR���Pvu�����ø����ø�л��գ�
�ٸ������Ƭ��������ォTac�����Ӻ�mCherryƬ�ν���PCR�����������Infusion�Լ�������pBBR1MCS2�Ǽܡ�tac�����Ӻ�mCherryƬ�Ρ����ö��ڷ����������Ӳ���ת��˾�����̬�У�ͨ������PCR�������Ծ��䣬˫ø����֤������ȡ���Ծ�����в�����֤��
1.2.4 ũ�˾�ϸ����Ĥ�����ԵIJⶨ
��1.2.1�½ڷ�������ò��Ӵ���ATCC15834��EHA105ũ�˾��������¾���1��24?h��48?h��ֱ�ȡ������LIVE/DEAD BacLight�Լ��вⶨũ�˾�ϸ����Ĥ�����ԣ��ö��ø���Dzⶨ��ӫ��ֵ����������Ϊ485?nm���ⶨ���䲨��Ϊ530?nm��ӫ��ǿ�ȣ�Emission 1����ɫ����
���ּ�������Ϊ485?nm���䣬�ⶨ��������Ϊ630?nm��ӫ��ǿ�ȣ�Emission 2����ɫ���������Ʊ����ߣ��������Һ��ũ�˾��ı�Ĥ�����ʡ�
1.2.5 ũ�˾����̲�Ҷ��ϸ��֮�������ԵIJⶨ
���������������̲����ڲ�ͬ��Һ����Ⱦ10?min��Ȼ��ȡ����������ֽ���ɶ����Һ�����泯��ƽ���ڹ������������ϣ�22�氵����һ��ʱ���ȡ�������ڶ��������λ���Һ��D-PBS���У�ϴȥ����ũ�˾������ѡȡ3��Ҷ���Ʊ���ʱװƬ��
����1�����̲�ҶƬ���ں���EHA105-pBBR1MCS2-Tac-EGFP��ATCC15834-pBB R1MCS2-Tac-EGFP�ľ�Һ��������������������48?h��
����2�����̲�ҶƬ���ں���EHA105-pBBR1MCS2-Tac-mCherry��ATCC15834-pBBR1MCS2-Tac-mCherry�ľ�Һ����Һ�����ӣ���������������������24?h��ȡ��������ȾҺ��ϴ���飻
�ٷֱ��ú�EHA105-pBBR1M CS2-Tac-EGFP��ATCC15834-pBBR1MCS2-Tac-EGFP�ľ�Һ�ٴ���Ⱦ��
ʹ�ü���۽������ֱ�۲촦��1�ʹ���2�ĺ���ɫӫ��ũ�˾����̲�ϸ����Χ�ֲ������
1.2.6 ũ�˾�DNA����ȡ�뷴ת¼
�������EHA105-pBI121��ATCC15834-pBI121�����̲�ҶƬ���ھ�Һ����Ⱦ10?min��ȡ5?mL��Һ������ũ�˾�RNA��ȡ�����̲ݰ���1.2.5�½ڵķ������й����������µ�D-PBS��������������-20���������������������̲�Ҷ��ϸ���ϵ�ũ�˾�ϴ�ѣ�
�ռ�����Һ������ũ�˾�RNA��ȡ��
ÿ�������ظ�3�Ρ�����trizol-up������RNA��ȡ����ת¼����ȫʽ����������˾TransScript?II All-in-One First-Strand cDNA Synthesis SuperMix for qPCR��One-Step gDNA Removal���Լ�����ɣ�cDNA�����ڨC20�棬ϡ��50��ʹ�á�
1.2.7 ũ�˾������������
��ũ�˾�cDNAΪģ�壬ũ�˾�������Ϊ�ڲλ�����ȫʽ����������˾PerfectStartTMGreen qPCR SuperMix�Լ��жԲ��ֻ������ʵʱӫ�ⶨ��PCR��⣬�ⶨEHA105��ATCC15834���ֻ���ͻ��������������ݽ��ȡ3�μ����ظ�ƽ��ֵ��
1.2.8 ��ͬũ�˾��鵼���̲�˲ʱת�����̲��Ŵ�ת��
���̲�ҶƬ�ֱ���뺬��EHA105-pBI121��ATCC15834-pBI121��Һ�У�����1����ˮ������2�ú����ӵ���ȾҺ����Ⱦ5?min��22�氵����2?d��
������EHA105-pBI121��Һ�������̲�����ˮ��ϴ���飬����GUSȾҺ��37��ڰ���ҹ��������75%�Ҵ���Ҷ���أ�Ҷ������ȥ�����ա�
������ATCC15834-pBI121��Һ�������̲��ú�500?mg��L-1ͷ������Ƶ�MSҺ������������20?min����ˮ��ϴ2�飬����ֽ���ɺ�����1/2MS+300?mg��L-1����͡+300?mg��L-1ͷ������ƹ����������Ͻ����Ѿ�������ÿ7?dת��1�Σ�ֱ�������Ծ����γɣ�2�ܺ�ͳ����ֲ�巢״���ͻ����ʡ�
2.1 ����Ũ�ȶԾ��������ʵ�Ӱ��
ũ�˾��Ļ�������ӵ�Ũ�Ⱥ�����ʱ���йء��ɱ�1��֪�����ӶԸ���ũ�˾�LBA4404��EHA105��������������������Ũ�ȵ����߶����������ƣ�������ʱ���ӳ��������������½������ơ�LBA4404������3?hʱ�������ʾ�Ϊ��ֵ�������������Ų���Ũ�ȵ����߶����ߣ�˵����ʱ���Ӷ�LBA4404��ٽ����á������Ų���Ũ�ȵ����ߣ��ٽ�������С����3��9?hʱ��Σ����Ӷ�EHA105�����������Դ��ڶ�LBA4404�������ʣ�
EHA105��3��6?h�������������Ũ�Ⱥ�����ʱ�������أ���9?hʱ�������������½��������
��12?hʱ����ͬŨ�ȵIJ��Ӷ�LBA4404�������ʴ���EHA105��
�Է���ũ�˾�ATCC15834��K599�������������Ų���Ũ�ȵ����߶����ߣ�
��������ʱ����ӳ������ߺͣ���ATCC15834��������������9?h��ʼ�½�������K599������6?h��ATCC15834�������ʾ����ڶ�K599�������ʡ�

��1 ���Ӷ�ũ�˾������ʵ�Ӱ��
ע��ͬһ��������ͬʱ���µıȽϣ���ͬСд��ĸ��ʾ0.05��������
Note: Comparison of the same strain at same time, different lowercase letters indicate significant difference (<0.05)
2.2 ��ͬũ�˾��Բ��������ԱȽ�
ǰ�����鷢�֣����о��е�4��ũ�˾��ڲ�ͬŨ�ȵIJ���������������60?hʱ��������ƽ̨�ڣ�ϸ�����������������ȶ������ѡȡ����ʱ��Ϊ60?h��ũ�˾��ⶨ���ڲ��Ӳ�ͬŨ���µ������ԡ���ͼ1��֪���ڲ�ͬŨ�Ȳ��Ӵ����£���ͬ�������������������LBA4404��ҺŨ�������Ũ�ȵ����ߣ�����仯��Խ�С���Բ�����������ǿ��ATCC15834��200?mg��L-1���Ӵ����£���ҺŨ�������������������飻
��������Ũ��Ϊ400��800?mg��L-1ʱ����ҺŨ�ȱ仯���첻������
K599��EHA105��1?000?mg��L-1���Ӵ����¾�ҺŨ���������ڶ���������������顣���Ͻ��������LBA4404��K599�Բ����н�ǿ������������EHA105��ATCC15834�Բ��ӽ�Ϊ���С���ˣ�ѡȡATCC15834��EHA105���к������顣
2.3 ���Ӷ�ũ�˾�ϸ����Ĥ�����Ե�Ӱ��
�ⶨATCC15834��EHA105������̡���ɫӫ��ֵ������ũ�˾��ı�Ĥ�����ʣ����������ũ�˾�ϸ����Ĥ�������������Ũ�Ⱥ;���ʱ��߶���ء�
��ͼ2��ʾ������1?hʱ��ATCC15834�ı�Ĥ����������������������죻
EHA105��Ĥ��������200��400?mg��L-1��600?mg��L-1�����������������������졣����24?h��ATCC15834��EHA105�Ķ����鱻Ĥ�������������ڴ����飬��ATCC15834�ı�Ĥ�����������Ũ�����߶����ͣ�EHA105����������ߺ͵����ơ�����48?hʱ��ATCC15834�Ķ�����ʹ����鱻Ĥ�����ʾ�������ͣ�
��������ڴ������Ҵ�����ı�Ĥ�����������Ũ�����߶����ͣ�����������
EHA105�Ķ������봦���鱻Ĥ������Ҳ����½����������������ڴ����顣

ע��ͬһ�����ڲ�ͬ����Ũ��֮��ıȽϣ���ͬСд��ĸ��ʾP<0.05��������

ע��ͬһʱ�䲻ͬСд��ĸ��ʾP<0.05������������ͬ
2.4 ���Ӷ�ũ�˾����̲�Ҷ��ϸ�������Ե�Ӱ��
����۽����������ͼ3��������Ũ�ȶ�ũ�˾��̲�Ҷ��ϸ����������Ӱ��������������48?hʱ������1����ATCC15834��EHA105��ȣ���ɫӫ����࣬˵�������̲�ϸ�����⸽�������ߣ�
ATCC15834��EHA105����ɫӫ�����Ų���Ũ�ȵ����߶����٣�˵���������������Ų���Ũ�ȵ����߶����͡�
ATCC15834��EHA105������24?h��������Ӻ�������24?h������2������ͼ3��ʾ��ũ�˾��������������̲�ϸ�����ϣ���ɫӫ�⣩��
���Ų���Ũ�����ߣ���ɫӫ����٣���ȥ�����Ӻ�ũ�˾��������������̲�ϸ�����⣬��ũ�˾�����������ɫӫ�⣩�������ȳ��ֳ����ߵ�����������
2.5 ���Ӷ�chv��������Ӱ��
��ͼ4��ʾ��ATCC15834�����Ӵ���1?h�������Ũ�ȵ����߱��������ߣ������ͱ��������������ߺ͵����ƣ��������������½������������ƣ�
����������800?mg��L-1���Ӵ����г�����ߵı����������������ڶ����飻
�����д������еı����������ڶ����飬���ڲ�������Ũ��Ϊ400?mg��L-1ʱ��������ߣ�
�������д������еı������������ڶ����顣������24?h����200?mg��L-1���Ӵ����µġ����ͱ������������ڶ���������������飬400��600?mg��L-1��800?mg��L-1���Ӵ����µġ����ͱ��������첻������
�������ڲ�������Ũ��Ϊ800?mg��L-1ʱ�ﵽ��ߣ�600?mg��L-1��1?000?mg��L-1���Ӵ������������������飻
������1?000?mg��L-1���Ӵ�����������������ڶ����飬������������������������������顣
EHA105�����Ӵ���1?h�͵ı��������ڶ����飻
��400?mg��L-1���Ӵ����£�����������������ڶ���������������飻
��200?mg��L-1��400?mg��L-1���Ӵ����£����������Ҳ�������ڶ���������������顣������24?h�������Ũ�����ߣ��͵�7����������������ߺͣ�����200?mg��L-1���Ӵ����£��������ﵽ���ֵ������400?mg��L-1���Ӵ����±������ﵽ���ֵ��
800?mg��L-1��1?000?mg��L-1���Ӵ����£��������������첻���������ͻ���ı������������ڶ����顣
���������������ͬ����ͬһ����Բ��ӵķ�Ӧ��ͬ��ͬһ���겻ͬ����Բ��ӵķ�ӦҲ��ͬ���ұ���仯��ʱ��Ͳ���Ũ�ȳ��ֳ���������ԡ�

ͼ3 ���Ӷ�EHA105��ATCC15834���̲�ҶƬϸ��֮�������Ե�Ӱ��

ͼ4 ���Ӷ�ũ�˾�EHA105��ATCC15834 chv�����������Ӱ��
2.6 ���Ӷ�vir��������Ӱ��
��ͼ5��ʾ��EHA105�����Ӵ���1?h�������Ũ�����ߣ��������д�����ı��������������ڶ����飬���������������첻������
���������ͳ������������ƣ����������ڶ����顣������24?h�������в��Ӵ������У��ͱ��������������ڶ����飬���������ڶ����顣
ATCC15834�У���Һ�����Ӵ���1?h���Ӵ����顢�����������ͻ�����������ڶ����飬�ͱ������������ڶ����顣������24?h���Ӵ������С��������������ͱ��������ڶ����飬���������ڶ����飻
��200?mg��L-1���Ӵ������У���������������������飻
��800?mg��L-1���Ӵ������б������������ڶ����飬�����������������ڶ����飻
��800?mg��L-1������������������ڶ����飬���������������еı����������ڶ����飨ͼ6����
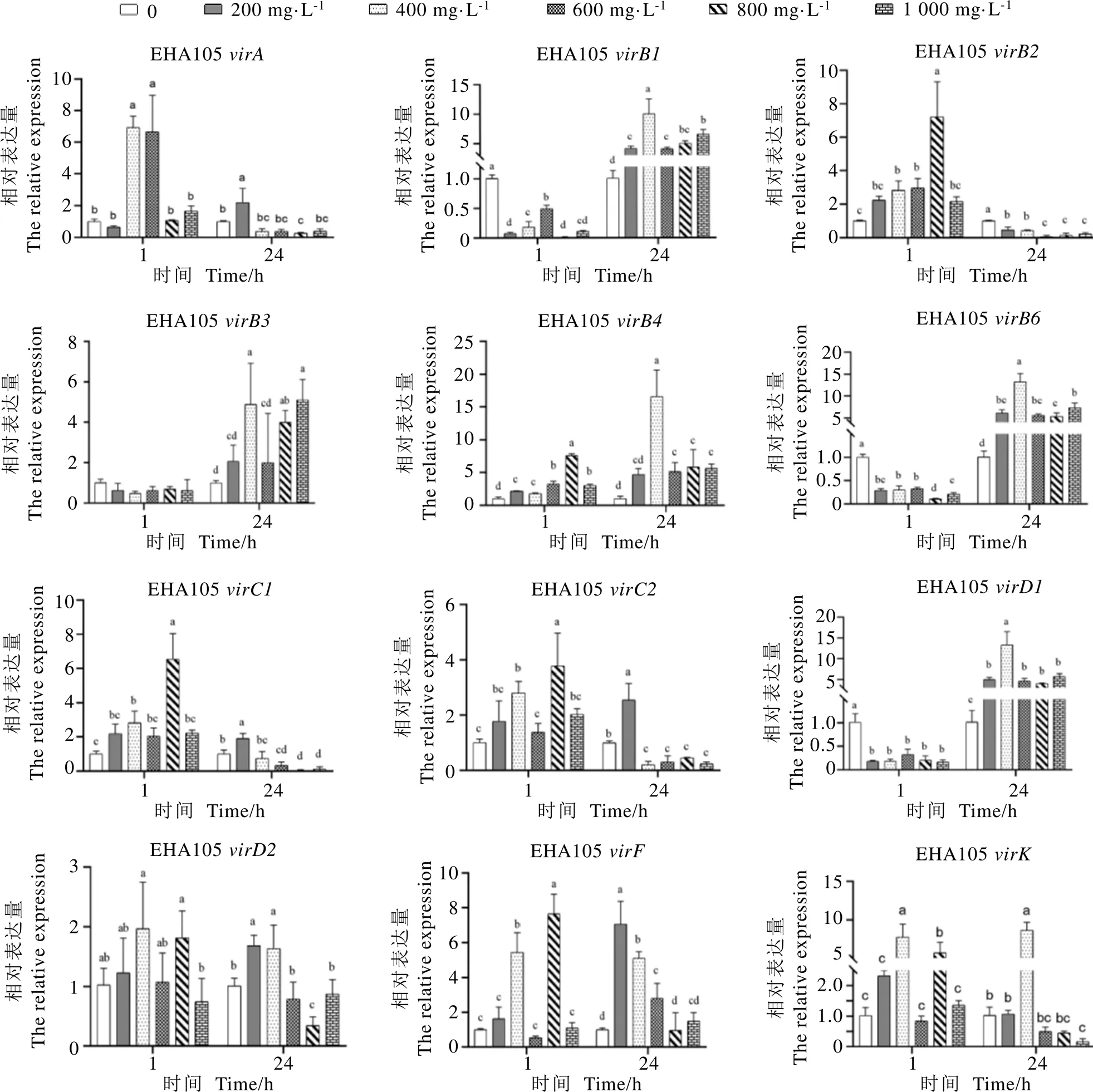
ͼ5 ���Ӷ�EHA105 vir�����������Ӱ��
2.7 EHA105��ATCC15834�鵼���̲�ת��
Ϊ̽�����Ӷ�ũ�˾��鵼ֲ���Ŵ�ת��Ч���Ƿ���Ӱ�죬ͨ��GUSȾɫ�����̲�ҶƬ˲ʱת��Ч������ͼ7��ʾ���ڹ�����3?d���Ӵ�������̲�Ҷ���пڴ����ֱֺ䣬�������Ũ�����ߣ��ֱ�̶ȼ��أ�����1?000?mg��L-1���Ӵ����³��ֻ�������������Ҷ�̱�����ɫ���ƻ���ֱ������
���о����ֺ���ͬŨ�Ȳ��ӵľ���Һ������Ⱦ�̲ݣ�GUSȾɫ���������ԡ����������ȣ������������Ũ�����ߣ�Ⱦɫ�̶ȱ�dz��
���ú���ͬŨ�Ȳ��ӵľ���Һ��Ⱦ�̲�Ҷ�̣���������ת�������ص�1/2 MS���������յ���״������ͳ��2�ܺ�ķ�״���յ��ʺͻ����ʡ�������2��ʾ��ATCC15834��״���յ��������Ũ�����߶����ͣ��������������Ũ�����߶����ӣ�
��1?000?mg��L-1���Ӵ��������յ��ʽ�Ϊ13.85%�������ʴ�33.85%��

ͼ7 ���Ӷ�ũ�˾�EHA105�鵼���̲�˲ʱת��GUSȾɫ��Ӱ��
���о������˲�ͬ����Ũ�ȶ�4��������־��ʣ���ѡ�������Բ��ӽ�Ϊ���еľ���ATCC15834��EHA105�����Բ���Ũ��Ӱ����������Ļ��������������̽�֡�ǰ���о���������Ũ�ȶ�������ʿ���Ϊ��ѧ������������ũ�˾���ֲ��ϸ���ƶ���������ũ�˾��������ϵͳ�����о����֣�EHA105��K599�ڲ�������Ũ��Ϊ0��200?mg��L-1ʱ�����ܵ����ƣ���200?mg��L-1����ʱ�����ͷ�����������Ũ�Ȳ��ӵĽ�ֵ��һ�����ģ���ĸ����ͬ����Բ��ӵ����жȲ�ͬ������ƫ���������ġ����⣬�ڲ�ͬ����Ũ����ϸ���������ʻ��ܵ���ͬ�̶ȵ����ƣ�ϸ������������Ҳ�������죬ODֵҲ���ܵ������������ص�Ӱ�졣
ͨ�����Ӷ�ATCC15834��EHA105���걻Ĥ�����Ե��о����֣�ũ�˾��ı�Ĥ����������ӵ�Ũ�Ⱥ�����ʱ���йأ���ǰ���о����һ��[27-29]����ͬŨ�Ȳ��Ӵ����£�EHA105�仯���ɲ����ԣ�ATCC15834��Ϊ���У�������1?hʱ�������������������IJ��졣���ﱻĤ�ķ�Ӧ���ƽ�Ϊ���ӣ��漰�����ź����ӵĵ��ء���������ʿ���ͨ����ϸ����Ĥ���ϣ��ͷ�֬���ǴӶ��ƻ�ϸ��Ĥ�ṹ��������[28]�����о���ATCC15834��EHA105����ı�Ĥ�����Ա��ֳ��Բ��Ӳ�ͬ����Ӧ���Ʋ��Dz�ͬ����İ�����Dz����ԭ������ɵġ�
ũ�˾��鵼����ת��Ч����vir����Ӱ�졣vir���ײ���T-DNA�ļӹ���ת�ƺ����ϵȹ��̣�����������������йء�Sharma��[30]�ö���������ȡ������־����飬���ֶ����ػ����Ʊ�Ƥ�������������������̸˾���ӫ��ٵ������Ϳݲ�ѿ�߸˾�������������С�־�Ũ�Ȼ���������ϸ�����̺�����Ƥϸ��֮��������ԡ����о�֤ʵ�˲�������������ũ�˾����̲�ϸ�����������ã�����Ч�������Ũ�ȳ�����ء�ũ�˾�����ֲ��ϸ��ʱ����ؼ����õ���ϸ������Ķ���ǣ��Ȼ������Ӱ��ũ�˾���ֲ��ϸ���ĸ�����ʶ�𣬲��Ӵ���24?h��ATCC15834�к�EHA105�еı������ܵ��������ƣ���˿��������ڵı������-1,2�Ͼ��Ǻϳ��ܵ��谭������ũ�˾��������������͡�һ���棬�ڲ�ͬũ�˾�����֮��ı���ģʽ������ͬ���ᵼ�²�ͬ����Է������ʺ�����ϸ����֪�����в����ԣ�
��һ���棬�������ʵĺ�����Ե�Ũ�ȷ���������Գ��������[31]�������Ӵ�����ͬ�������������ʺ����ϵIJ��죬Ҳ��Ӱ��ũ�˾��Բ��������Բ����ԭ��֮һ��
�̲�ת����ϵ����ȾҺ������ӣ���ͬ�̶ȵ�����˲ʱת��Ч�ʺ��ȶ�ת��Ч�ʣ���״���յ��ʴ�ͣ��̲�ҶƬ�����˷��ơ��ֱ��֢״�������Ũ�����ߣ�������ֲ��ƻ��������ʲ�����������ֲ���������ܵ����ƣ������̲��Ŵ�ת��Ч���½������о�������ֲ��ϸ���ܵ����˺���Ҫ������Ⱦǰ����Ϊ����Ļ�е���˺ͺ�������ũ�˾����˺�����ֲ���ܵ��˺�������øϵͳ������ٽ��������ʵĺϳɺͻ��ۣ���ߵֿ�����������������ױ��������ø�������������ʣ���������ϸ����������������֯�ֱ䡢����[32]��
���о�ͨ������ũ�˾��Բ��������ԡ����Ӷ�ũ�˾�ϸ����Ĥ�����Ժ�ϸ�������ԡ�����ͻ�������Ӱ���Լ����ӶԸ���ũ�˾�EHA105�ͷ���ũ�˾�ATCC15834�鵼���̲�ת��Ч�ʵ�Ӱ�죬Ϊũ�˾��鵼�IJ����Ŵ�ת����ϵ�ṩ��һ�������ۻ�����Ϊ��һ����߲����Ŵ�ת��Ч�����������̽�֡�
[1] �ڼ���, �����, ���, ��. �Ͼ�軨���ضԡ����ߡ���Ӱ�켰�俪������[J]. ����ũҵ��ѧ, 2018, 46(10): 39-41.
Tong J Y, Yang Y J, Yang F H, et al. Influence of anthocyanins of Zijuan tea on hypertension, hyperlipidemia and hyperglycemia and its development and utilization [J]. Journal of Anhui Agricultural Sciences, 2018, 46(10): 39-41.
[2] ������. ���ӵĹ�Ч����ȡ�����о���չ[J]. ����Ƽ�, 2011, 27(15): 57-58.
Pan W C. Research Progress on the efficacy and extraction technology of tea polyphenols [J]. Gansu Science and Technology, 2011, 27(15): 57-58.
[3] Cabrera C, Gimenez R, Lopez M C. Determination of tea components with antioxidant activity [J]. Journal of Agricultural and Food Chemistry, 2003, 51(15): 4427-4435.
[4] Chan E W C, Soh E Y, Tie P P, et al. Antioxidant and antibacterial properties of green, black, and herbal teas of[J]. Pharmacognosy Research, 2011, 3(4): 266-272.
[5] Souissi M, Ben Lagha A, Chaieb K, et al. Effect of a berry polyphenolic fraction on biofilm formation, adherence properties and gene expression of streptococcus mutans and its biocompatibility with oral epithelial cells [J]. Antibiotics, 2021, 10(1): 46. doi: 10.3390/antibiotics10010046.
[6] �����. ���ӵ���ȡ���־��������־������о�[D]. �ɶ�: ������ѧ, 2017.
Li K X. Study on extraction, bacteriostasis and bacteriostatic mechanism of tea polyphenols [D]. Chengdu: Xihua University, 2017.
[7] Bhattacharya A, Sood P, Citovsky V. The roles of plant phenolics in defence and communication duringandinfection [J]. Molecular Plant Pathology, 2010, 11(5): 705-719.
[8] Matsumoto S, Fukui M. Effect of acetosyringone application on-mediated gene transfer in tea plant () [J]. Bulletin of the National Research Institute of Vegetables, Ornamental Plants and Tea (Japan), 1999(14): 9-15.
[9] Mondal T, Bhattacharya A, Ahuja P, et al. Transgenic tea [(L.) O. Kuntze cv. Kangra Jat] plants obtained by-mediated transformation of somatic embryos [J]. Plant Cell Reports, 2001, 20(8): 712-720.
[10] ������, ������, ���. ũ�˾��鵼�������ǹ����������ֲ���Ŵ�ת���ϵ�Ӧ��[J]. ����ֲ��ѧ��, 2005, 25(1): 205-210.
Geng L Z, Liu C L, Li F G. Application of the combination of-mediated transformation and particle bombardment in plant genetic transformation [J]. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(1): 205-210.
[11] �ź�ƽ. ��ת������ϵ���Ż��Ϳ���ή�������ת��[D]. ����: �㽭��ѧ, 2008.
Zhang H P. Optimization of cotton transformation system and the transformation of resistant gene to[D]. Hangzhou: Zhejiang University, 2008.
[12] ��ˮƽ. Ұ��������³��ķ��ή��������ػ����ܼ���[D]. �Ͼ�: �Ͼ�ũҵ��ѧ, 2011.
Li S P. Functional analysis ofgene related toresistance [D]. Nanjing: Nanjing Agricultural University, 2011.
[13] Ҷ�˹�, ����, �����, ��. С��ת������������[J]. �Ŵ�, 2011, 33(5): 422-430.
Ye X G, Chen M, Du L P, et al. Description and evaluation of transformation approaches used in wheat [J]. Hereditas, 2011, 33(5): 422-430.
[14] ���г�, ���, ʩ��, ��. ����ˮ��������ũ�˾�������������źŷ���[J]. ��ѧͨ��, 1995, 40(12): 1126-1128.
Yin Z C, Li B J, Shi J, et al. There are signaling molecules for genes in the toxic region of novelin rice [J]. Chinese Science Bulletin, 1995, 40 (12): 1126-1128.
[15] Bhattacharya A, Sood P, Citovsky V. The roles of plant phenolics in defence and communication duringandinfection [J]. Molecular Plant Pathology, 2010, 11(5): 705-719.
[16] Cabrera C, Gimenez R, Lopez M C. Determination of tea components with antioxidant activity [J]. Journal of Agricultural and Food Chemistry, 2003, 51(15): 4427-4435.
[17] Chan E W C, Soh E Y, Tie P P, et al. Antioxidant and antibacterial properties of green, black, and herbal teas of[J]. Pharmacognosy Research, 2011, 3(4): 266-272.
[18] ���, ��С��, �����, ��. ���ӵĿ��������о���չ[J]. ʳƷ��ҵ�Ƽ�, 2015, 36(21): 385-389.
Yang H L, Liu X X, Zhu J L, et al. Research progress in antibacterial properties of tea polyphenols [J]. Science and Technology of Food Industry, 2015, 36(21): 385-389.
[19] Sharma A, Gupta S, Sarethy I P, et al. Green tea extract: possible mechanism and antibacterial activity on skin pathogens [J]. Food Chemistry, 2012, 135(2): 672-675.
[20] ���, ������, ������, ��. �����̲�����־����Լ�������о�[J]. �Ĵ�ũҵ��ѧѧ��, 2019, 37(6): 821-827.
Chen C, Xu Y M, Lin B B, et al. Antibacterial activity and mechanism of green tea polyphenols from qinling mountains [J]. Journal of Sichuan Agricultural University, 2019, 37(6): 821-827.
[21] Gonzalez-Rivera C, Khara P, Awad D, et al. Two pKM101-encoded proteins, the pilus-tip protein TraC and Pep, assemble on the Escherichia coli cell surface as adhesins required for efficient conjugative DNA transfer [J]. Molecular Microbiology, 2019, 111(1): 96-117.
[22] Matthysse A G. Attachment ofto plant surfaces [J]. Frontiers in Plant Science, 2014, 5: 252. doi: 10.3389/fpls.2014.00252.
[23] ������. ֲ����֬���ۻ�����Ʒ�ʸ���: 1.����RNAi��������֬Ʒ��;2.�ĺ�ľ��������֬����������[D]. ����: �й���ѧԺֲ���о���, 2007.
Wang G L. Vegetable oil accumulation mechanism and quality improvement: 1. The improvement of soybean oil quafity using RNAi; 2. Triacylglycerols in stems of[D]. Beijing:Institute of Botany, the Chinese Academy of Sciences, 2007.
[24] �δ���. �����ض�ũ�˾��鵼�IJ����Ŵ�ת����Ӱ��[D]. �Ϸ�: ����ũҵ��ѧ, 2014.
Song D P. Effects of catechins on-mediated gene transformation of[D]. Hefei: Anhui Agricultural University, 2014.
[25] ë����, ʩ����, ����, ��. ��Ҷ�����ضԷ���ũ�˾����������ü����Ӿ���ɸѡ�о�[J]. ��Ҷ��ѧ, 2007, 27(3): 243-247.
Mao Q L, Shi Z P, Li L, et al. Study on inhibitation ofby tea catechin and screening of anti-polyphenol strain [J]. Journal of Tea Science, 2007, 27(3): 243-247.
[26] ��÷��, �ܽ�, �ɺ�, ��. ����ũ�˾����ͷ����ܱȽ���ѱ���о�[J]. ��Ҷ��ѧ, 2008, 28(3): 189-194.
Cang M Q, Zhou J, Cheng H, et al. Studies on polyphenol-resisting characteristics of several kinds ofand the demestication of polyphenol-resisting strain [J]. Journal of Tea Science, 2008, 28(3): 189-194.
[27] Alshuniaber M A, Krishnamoorthy R, Alqhtani W H. Antimicrobial activity of polyphenolic compounds fromagainst food-borne bacterial pathogens [J]. Saudi Journal of Biological Sciences, 2021, 28(1):459-464.
[28] Kuo C Y, Zupko I, Chang F R, et al. Dietary flavonoid derivatives enhance chemotherapeutic effect by inhibiting the DNA damage response pathway [J]. Toxicology and Applied Pharmacology, 2016, 311: 99-105.
[29] ����.sp.QY101������ǵķ��봿������ϸ�����ﱻĤ�����о�[D]. ɽ��: �й������ѧ, 2011.
Jang P. The studies on isolation, purification and antibiofilm activities of the exopolysaccharide fromsp. QY101 [D]. Shandong: Ocean University of China, 2011.
[30] Sharma A, Gupta S, Sarethy I P, et al. Green tea extract: possible mechanism and antibacterial activity on skin pathogens [J]. Food Chemistry, 2012, 135(2): 672-675.
[31] ����ƽ, ����, ʦ־��, ��. ũ�˾�T-DNA������ػ����о���չ[J]. ����ֲ������, 2017, 15(5): 1752-1761.
Wang G P, Zhang T, Shi Z G, et al. Research progress on T-DNA transfer related genes mediated of[J]. Molecular Plant Breeding, 2017, 15(5): 1752-1761.
[32] ����. ���˻�ҩ������֯�յ����ֻ������Ϳ��Ʒ����о�[D]. �ɶ�: �Ĵ�ũҵ��ѧ, 2016.
Li H. Studies on callus induction and mechanism of browning and control methods in loquat [D]. Chengdu: Sichuan Agricultural University, 2016.
Effects of Tea Polyphenols on-mediated Plant Genetic Transformation System
LI Jing1,3,4,LIN Cairong1,3,4,HUANG Yan2,3,4,DENG Xuming1,3,4,WANG Yiqing1,3,4,SUN Weijang1,3,4*
1. College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 2. Anxi College of Tea Science, Fujian Agriculture and Forestry University, Quanzhou 362400, China; 3. Fujian Tea Industry Engineering Technology Research Center, Fuzhou 350002, China; 4. Ministerial and Provincial Joint Innovation Centre for Safety Production of Cross-Strait Crops, Fujian Agriculture and Forestry University, Fuzhou 350002, China
In order to study the effect of tea polyphenols on the infection efficiency ofin tea plant genetic system mediated by, fourstrains LBA4404, EHA105, ATCC15834 and K599 were used as research objects. The phenolic resistance, membrane integrity, adsorption,gene andgene expression and genetic transformation ofunder different concentrations of tea polyphenols were analyzed. The results show that: (1) the phenolic resistance of the four strains followed the order of LBA4404>K599>EHA105>ATCC15834, and EHA105 and ATCC15834 were more sensitive to tea polyphenols. (2) The ratio of ATCC15834 with intact membrane was negatively correlated with the concentration of tea polyphenols and the standing time, and the lowest intact membrane rate of EHA105 appeared 48?h after treatment with 600?mg��L-1tea polyphenols. (3) The adsorption capacities of ATCC15834 and EHA105 on tobacco mesophyll cells decreased with the increase of tea polyphenol concentrations. After treatment with tea polyphenols for 24?h, the expressions ofandin ATCC15834 and EHA105 were significantly inhibited. The expression ofwas positively correlated with the sensitivity to low concentration phenols. (4) After infecting tobacco by thesolution (containing tea polyphenols), the transient and stable transformation efficiencies were inhibited to varying degrees, and the induction rate of hairy roots was greatly reduced. When the concentration of tea polyphenols was 1?000?mg��L-1, ATCC15834 achieved the lowest root induction rate of 13.85% and the highest necrosis rate of 33.85%. In conclusion, tea polyphenols could reduce the activity and membrane integrity rate of, and reduce the expression ofgene to affect its adsorption, which finally suppress the instantaneous conversion efficiency and the stable conversion efficiency in tobacco conversion system to varying degrees.
tea polyphenol,-mediated, adsorbability,andgene, genetic transformation
S571.1
A
1000-369X(2022)04-477-14
2021-12-30
2022-03-29
������Ȼ��ѧ����31770732��������ʡ��У��ѧ������Ŀ��2019N5007����������Ҫ����ԭ������Դ�����ϵ�о���K1520005A04�����������츣��Ҷ��չ�����Ƽ����»���FJZTF01��
���Ů��˶ʿ�о�������Ҫ���²����������������\���о���*ͨ�����ߣ�swj8103@126.com
����ϲ�� ��Һ�����ʲ��� ˫øˮ�����۵����Ʊ�ACE�����ĵĹ����Ż��о��ִ�ũ��Ƽ�(2022��1��)2022-01-21�߲�ʹ�ò�ͬ������ʽ��ũҩ����Ӱ����о�ʳƷ��(2021��7��)2021-07-19���ϸ������ֲҵ�ϵ�Ӧ���о���չ����ũҵ��ѧ(2020��8��)2020-06-01��Чѡ��ɳ���Ͼ����������ݿ�ѧ(2020��3��)2020-05-13����ò��Ӹ�������ƷӪ�������ر����°���(2019��8��)2019-08-30�Ȳ��ܹ��ͼ��ʣ�����������ͥ���ഺ(2019��1��)2019-01-25�����������������������ͥ��������(2017��11��)2017-11-08�̹��ϼ״���ȡ���ֲ�ﻯ�����õ��о��Ӳ�ѧ��(2016��4��)2017-05-31����֬��ȡ��Իƹ�̿�Ҳ��������ƻ��Խ���ũҵ��ѧ(2016��1��)2017-05-17�ٵ�����HYS������Ͳ����ӷ�ѿ��Ӱ�����ũҵ��ѧ(2017��7��)2017-05-13